Anatomy and Physiology: Connecting with Synapses
Connecting with Synapses
Synapses have all the fun. The axons and dendrites just carry the message, but it is the ability for these cells to actually talk to one another that is the true glory of nervous tissue! The talking takes place at the synapses, between axons and dendrites, axons and cell bodies, or axons and effectors (such as muscles). In the case of neuron to neuron synapses, the first cell is the presynaptic neuron (with its presynaptic membrane), and the receiving cell is the postsynaptic cell (with its postsynaptic membrane).
There are actually two types of synapses: electrical and chemical. Electrical synapses are also called gap junctions and are formed by protein channels that physically connect two cells. These protein channels, called connexons (because of the way they connect cells), speed up the transport of ions from cell to cell, so that a wave of depolarization can travel very quickly, for example, from one cardiac cell to another, making multiple cells act like one.
Chemical synapses, however, are the basis of the vast majority of synaptic connections in the CNS and PNS. The majority of synapses are called cholinergic synapses, due to their use of acetylcholine (ACh) as the neurotransmitter (the chemical that sends the message from cell to cell, initiating a depolarization of the second cell. For a detailed run-down of this process, see the discussion of the synapse between a motor neuron and a muscle cell in The Structure of the Muscles and Muscle Cells.
One aspect of acetylcholine that is not discussed in The Structure of the Muscles and Muscle Cells is that of the recycling in the synapse. I mention the enzyme acetylcholinesterase (ACh-esterase) in the synaptic cleft. ACh-esterase breaks ACh into acetate (A) and choline (Ch). The acetate is absorbed and used by cells in the area. The choline, however, is absorbed by the synaptic end bulb. Coenzyme A (CoA) is taken up by mitochondria there, and converted into acetyl-CoA. The acetyl-CoA then combines with the choline to produce acetylcholine (ACh) and a coenzyme A (CoA), and the cycle continues.
Neurotransmitters
Medical Records
Physical addiction is based upon the building of extra receptors in the brain. The body, in its effort to maintain homeostasis, will build extra receptors (for example, endorphin receptors) to deal with the influx of the drug (for example, heroin). Over time, the addict will need to take more of the drug to get the same high; this phenomenon, caused by the extra receptors, is called tolerance. Withdrawal is hard due to the large number of now empty receptors. In the case of heroin withdrawal, since endorphins block pain, many empty recep-tors make quitting cold turkey very painful.
The field of neuroscience continues to find new neurotransmitters. Despite many of these discoveries, it turns out that the action triggered by the neurotransmitter has to do with the receptor, rather than the neurotransmitter, for the same chemical may stimulate certain neurons, but inhibit others. This leads to the idea of excitatory neurotransmitters, which initiate depolarization of the postsynaptic membrane, and inhibitory neurotransmitters, which causes hyperpolarization, thus inhibiting the depolarization of the postsynaptic membrane.
Some of these many neurotransmitters have a direct effect, as seen with ACh, but others trigger a response indirectly, using a secondary messenger not unlike that seen in the case of protein hormones (see Hormones). In addition to ACh, some neurotransmitters may be biogenic amines (epinephrine, norepinephrine, histamine, dopamine, and serotonin), amino acids (glutamine and aspartate), hormones (ADH, oxytocin, and so on), ATP, or even dissolved gases (carbon monoxide and nitric oxide). One interesting group of neurotransmitters is called neuropeptides, which may be only a few amino acids long; the most famous of these is called endorphin or enkephalin. Endorphin is short for endogenous morphine, or the body's own morphine, and it is the endorphin receptors to which opium, morphine, codeine, and heroin bind.
Types of Neurons
In a functional sense, neurons are classified according to the direction of their nerve impulse: afferent (or sensory) neurons travel to the brain, and efferent (or motor neurons, stimulating effectors such as muscle) travel from the brain. In addition, the majority of neurons in the body actually carry messages from one neuron to another; these middle men are called interneurons. Some neurons function by stimulating the following neuron, while others function by inhibiting the following neuron. Lastly, neurons also vary in terms of the layout of the dendrites and axon (see Figure 19.4).
One form is the multipolar neuron, in which there are multiple short dendrites, and one long axon; these neurons, which look rather like they have dreadlocks, are very common in the brain and spinal cord. Bipolar neurons, as their name suggests, have the cell body in the middle, with one long dendrite, and one long axon; all bipolar neurons are sensory. Unipolar neurons are also, and more accurately, called pseudounipolar neurons. These rather bizarre-looking neurons have a continuous track connecting the dendrite (which may be myelinated) to the axon, with the cell body off to the side, connected by a stalk; sensory neurons in the PNS mostly take this form. Anaxonic neurons really aren't without axons, but their axons and dendrites are short and similar in length and appearance; the function of these neurons is still under investigation. I discuss sensory neurons in more detail in The Central and Peripheral Nervous Systems.
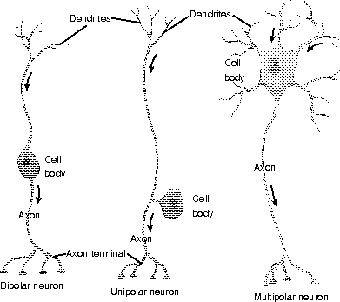
Figure 19.4Neuronal types. (LifeART©1989-2001, Lippincott Williams & Wilkins)
Types of Circuits
The connection between neurons leads to some interesting connections called circuits. The nature of the arrangement of each type of circuit says a great deal about the function of that circuit. Series circuits, the least interesting of the bunch, are merely a connection of single neurons in a line; such circuits basically just carry information from one center of the brain to another for processing.
On the other hand, circuits may either diverge (one neuron connecting to many) or converge (many neurons connecting to one). Their arrangement is an aspect of their function. A diverging circuit of sensory neurons, for example, would split in terms of the areas of the brain to which the sensory input goes. Visual images would need to respond to the amount of light to open or close the pupil, or to interpret words, or identify the emotions behind a facial expression; a diverging circuit would thus send the same signal to multiple locations in the brain.
Converging circuits, on the other hand, would most likely be motor neurons. Consider your breathing. Chances are you haven't been thinking about that as you have been reading this section (or if you have, you should pay more attention to what you are reading).
In that sense, isn't breathing involuntary? If you said yes, you're right, but what about holding your breath? Isn't that voluntary? If you said yes, you are also right! The only way that is possible is through a converging circuit, which will allow stimulus for the diaphragm by either a voluntary or involuntary neuron. In the case of holding your breath, the voluntary neuron must have an inhibitory effect on the motor neuron, or else the involuntary command will take over. Imagine suddenly breathing while under water, and you'll get the idea of the importance of this inhibition!
Parallel circuits look similar to diverging circuits, and, indeed, they must first diverge before they can become parallel. Such circuits help explain responses to sudden painful stimulus. Have you ever picked up something that was way too hot to hold? Think about what you did. Did you drop it suddenly? Did you also pull your arm back? What about your voice? Did you suddenly scream out a bunch of syllables my editor won't let me put in this section? All of those sudden responses to that stimulus were sent to different parts of the brain, and then to different effectors, through a parallel circuit.
Reverberating circuits differ from the others in that they have neurons with branches that go backward to stimulate the circuit earlier in the chain. Remember that neurons only send messages one way, so such a circuit really requires a place for an axon to backtrack to a dendrite of an earlier axon. But why would the body do such a thing. Once again, such a circuit implies a particular function. Such a circuit, for example, would be used to control a particular repetitive action, such as breathing. In higher levels of the brain, reverberating circuits control consciousness, for without a constant restimulation through the circuit, there would be no continuity to our consciousness. Such circuits require specific circumstances to break the circuit, as when we fall asleep!
The Parts of an Arc
A simple way to explore the function of neurons, both alone, and in concert, is to look at a reflex arc. To understand the parts of a reflex arc, it is helpful to think about the spinal cord, the dorsal and ventral roots to the spinal nerve, and feedback loops. Reflexes follow the basic pattern of a negative feedback loop, with a receptor, a control center, and an effector. Given that the purpose of a reflex is to reverse a stimulus, in other words negative feedback, reflexes help to maintain homeostasis.
Let's take a look at a relatively simple polysynaptic reflex, shown in Figure 19.5, for it will illustrate the concept well. First of all a stimulus, such as stepping on a tack, triggers pain receptors, thus initiating an impulse in a sensory (afferent) neuron. That impulse travels through the dorsal root ganglion to the dorsal root to the spinal cord. The posterior gray horn forms synaptic connections with two interneurons, which, as the name implies, are not the end of the line. A monosynaptic reflex, on the other hand, forms a single synapse with a motor (efferent) neuron.
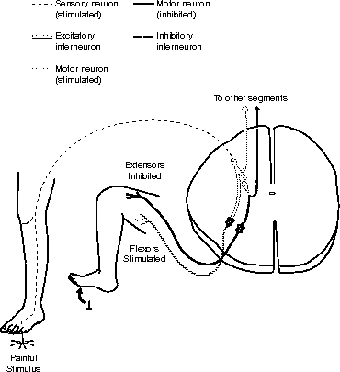
Figure 19.5This polysynaptic reflex has not only a sensory neuron, but two interneurons (one to triggering a neuron that stimulates the flexors, and one that triggers a neuron that inhibits the extensors).
Although these interneurons may send messages to other segments of the spinal cord, or to the brain, here I am going to consider the synapses to motor neurons. It is important to consider here that muscles, our effectors, come in antagonistic pairs (see The Structure of the Muscles and Muscle Cells and The Muscles); as one muscle contracts, its antagonist relaxes, and vice versa. If both muscles contract there would be no movement! Therefore, one interneuron is called the excitatory interneuron, because it stimulates a motor neuron, and the other is called an inhibitory interneuron, because it inhibits a motor neuron.

Excerpted from The Complete Idiot's Guide to Anatomy and Physiology © 2004 by Michael J. Vieira Lazaroff. All rights reserved including the right of reproduction in whole or in part in any form. Used by arrangement with Alpha Books, a member of Penguin Group (USA) Inc.
To order this book direct from the publisher, visit the Penguin USA website or call 1-800-253-6476. You can also purchase this book at Amazon.com and Barnes & Noble.
Trending
Here are the facts and trivia that people are buzzing about.





