Genetic Variation and Natural Selection: Post-Mendelian Inheritance Factors
Post-Mendelian Inheritance Factors
Recall from Inheritance that most animals, including you, are diploid, meaning that each of your traits is controlled by the interaction of at least two genes. Mendel's work concentrated on the effect of a single gene from the mother and a single gene from the father to determine the genotype and phenotype of the offspring based on dominant and recessive genes. We now know that often multiple genes are involved in strange and intriguing ways to influence the phenotype of the offspring. The next section focuses on seven such mechanisms.
Bionote
More than 200 human traits are controlled by a single gene pair, such as dwarfism, cataracts, cystic fibrosis, and albinism.
Incomplete Dominance
Whenever two genes combine and the offspring's phenotype is a compromise between the effects of the two genes, then neither gene has expressed dominance over the other gene. In fact, one gene is incompletely dominant over the other gene that is inactive. Mendel's work with peas never encountered incomplete dominance because the heterozygous genotype always expressed the dominant gene in the phenotype.
The active gene is apparently not strong enough to compensate for the loss of effect by the inactive allele. The classic example of incomplete dominance is the cross between a red snapdragon and a white snapdragon. According to Mendel, the offspring should be either red or white. In reality they are all pink, indicating that neither allele is completely dominant but one is incompletely dominant. Therefore, a weak expression of one allele, the red one, is actually expressed in a heterozygous offspring.
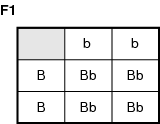
Incomplete dominance becomes evident in the F2 generation as detailed in the following Punnet square analysis of a fictitious trait: B = blue, b = yellow. A cross between a blue (BB) parent and a yellow (bb) parent, BB × bb, should give all blue offspring in the F1 generation. Refer to the illustration Parents: BB × bb.
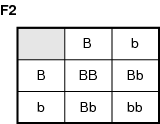
Instead, the Bb genotype is green, a blending of the two colors. This indicates that the B gene is not strong enough to dominate the effect of the b allele completely. If two heterozygous individuals are mated, the results are even more intriguing. Refer to the illustration Parents: Bb × Bb.
The genotypes are BB (1); Bb (2); bb (1). A 1:2:1 ratio is typical of Mendelian inheritance patterns. However, the phenotypic ratio is: blue (1), green (2), yellow (1). This 1:2:1 ratio is not expected by Mendel's predictions.
Codominance
Codominance is a type of allele dominance that occurs when both alleles are active and expressed, as sometimes happens in a heterozygous individual. This phenomenon is most commonly displayed in the roan coat color of horses and cows. A roan color is actually the result of a codominance by a gene for red hair and a gene for white hair. In a roan-colored animal, there are equal numbers of both colors evenly dispersed within the coat, creating a ghostly brown color. Another example is Erminette chickens. They are a mottled black and white color because they have roughly equal numbers of black and white feathers. In their case, a gene for white feathers and a gene for black feathers are codominant in the heterozygous individuals.
Multiple Alleles
Although individual humans carry a maximum of two alleles that control the expression of a trait, often there are more than two different alleles in a given population that code for the same trait. When three or more alleles are available for a particular gene, the inheritance of those alleles follows a multiple alleles pattern. The most common case for demonstrating multiple alleles is the inheritance of human blood types: A, B, AB, and O. Combinations of three possible alleles give rise to the four blood types. Because you can only have two alleles for any gene, not everyone carries every type of allele available; but within a population, all of the multiple alleles are present. Sexual reproduction within that population keeps the alleles available for future generations and continues the genetic variation.
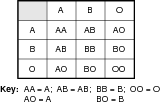
An expanded Punnet square shows the possible blood type combinations that exist in the human population. Refer to the illustration Human population blood type combinations.
Remember although there are three possible alleles listed on each axis, a human can only possess two of these. However, other individuals in the human population may carry a different set of two alleles, so that all three types are present and active in the human population.
Linked Genes
Each chromosome normally carries thousands of genes. When certain neighboring genes on the same chromosome are inherited together, they are considered . Linked genes are an anomaly in Mendel's theory of independent assortment because they tend to remain together as a segment and therefore function as a cluster of genes rather than individual genes. In humans, the autosomal genes for red hair, freckles, and a fair complexion are linked genes.
Sex-Linked, or X-Linked Genes
The linkage in sex-linked traits is completely different from linked genes. A sex-linked trait is one whose allele is located on one of the sex chromosomes, usually the X for autosomal traits. Most sex-linked genes are recessive and are only expressed in homozygous females and males. Because males normally inherit one X chromosome from the mother only, the recessive traits found on the X chromosome are passed from mother to son. It is more difficult to pass an X-linked trait to a daughter because she inherits an X from both parents, so both parents would have to possess and pass on the recessive gene. The recessive gene on the X chromosome is expressed in males because it is the only gene present. In females, it can be hidden by the presence of a dominant gene on the other X chromosome. In humans, sex-linked traits include colorblindness, albinisms, and hemophilia.
Refer to the illustration Colorblindness pedigree. A heterozygous “carrier” mates with a “carrier” female (Xc = gene for colorblindness). A carrier is an individual who is heterozygous for a condition but does not display the trait. Therefore, a carrier for colorblindness is not colorblind but can pass on the gene for colorblindness.
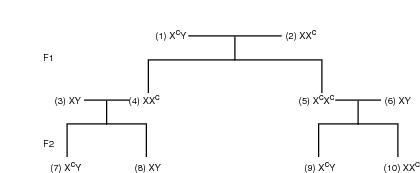
In the F1 generation, one colorblind female and one carrier female each mate with a noncolorblind male. In the F2 generation, individuals 7 and 8 are brothers, one is colorblind, the other is not. The colorblind male inherited the colorblind gene from the heterozygous carrier mother; the noncolorblind son inherited the normal X chromosome that did not contain the gene for colorblindness. Likewise, individual 9 could only receive a gene for colorblindness from his mother, number 5. The sister, number 10, is a heterozygous carrier.
Sex-Influenced Genes
Certain genes are activated by hormones secreted by the sex organs, so they display different phenotypes when found in males than they do when found in females, even though their genotypes are identical. The standard sex-influenced study in humans is male pattern baldness, which is influenced by testosterone and can affect both males and females. However, it is dominant in males and recessive in females! In the homozygous state, both males and females become bald; in the heterozygous genotype, females show no signs of baldness, but males become bald. Interestingly, sex-influenced genes are generally not found on the sex chromosomes, so both sexes can have the same genotype and different phenotypes!
Bionote
Polygenic inheritance is the opposite of pleiotropic inheritance, in which a single gene affects several characteristics.
Polygenic Inheritance
So far we have limited gene expression to those traits expressed by the action of one gene. In most primates, including you, traits are governed by the interaction of multiple genes. The genes of polygenic traits may be interspersed on the same chromosome or on completely different chromosomes. The additive effects of numerous genes on a single phenotype create a continuum of possible outcomes. A typical example is eye color in humans, which under normal genotypes is in varying shades of ebony brown to crystal blue, to Kelly green, and all points in between. Polygenic traits are also most susceptible to environmental influences. For instance, tallness is controlled by polygenes for skeleton height, but their effect may be retarded by malnutrition, injury, and disease.
Environmental Influences
Natural environmental influences on the expression of genotypes tend to be an advantage for the individual, whereas man-made environmental influences are both positive and negative for the individual. Natural environmental influences include the phenomenon of color change in the Arctic fox from red-brown in the summer months to pure white during the winter season for better camouflage. The genes that produce the red-brown summer pigment are blocked by cold temperatures, causing the hair to grow with no color (therefore, white). Another colorful example is the interaction between the color of the hydrangea flower, which is blue in acidic soils and pink in alkaline soils. A recent study also linked improved diet in infants and adolescents to a taller average height in the United States and Europe with the opposite effect in famine-stricken populations. The genetic complement of an individual is inherited; however, the environmental effect on these genes may alter their application and expression.
Excerpted from The Complete Idiot's Guide to Biology © 2004 by Glen E. Moulton, Ed.D.. All rights reserved including the right of reproduction in whole or in part in any form. Used by arrangement with Alpha Books, a member of Penguin Group (USA) Inc.
To order this book direct from the publisher, visit the Penguin USA website or call 1-800-253-6476. You can also purchase this book at Amazon.com and Barnes & Noble.
Trending
Here are the facts and trivia that people are buzzing about.





