Specialized Cell Structure and Function: Modifications and Adaptive Functions
Modifications and Adaptive Functions
As organisms felt the pressure of natural selection and attempted to colonize new territories, the need for advancements in both structure and function were necessary. They evolved with modified structures (evolution) or were created with the appropriate structures (creationism) to fit the environment. Advancements in structure and function created complex life-supporting systems that are more versatile and allow the organisms greater freedom for colonization in fringe territories. Photosynthesis, respiration, and protein synthesis are typical examples of complex chemical phenomena that occur around and within us constantly.
Location, Structure, Function
Bioterms
Photosynthesis is the process whereby plants convert light energy into chemical energy by making a variety of organic molecules, including glucose, from carbon dioxide (CO2) and water (H2O).
Most people think that photosynthesis is a process in which six carbon dioxide molecules react with six water molecules using the power of light to create one molecule of glucose and six molecules of oxygen. Although this simplified equation is fundamentally correct, it does not highlight the various reactions that make photosynthesis an interesting plant cellular event.
It is believed that 3 to 3.5 billion years ago, photosynthesis changed the environment of the early Earth from one that lacked oxygen to an oxygen-rich environment that, in turn, profoundly changed the type of early life that could live there and forced the extinction of many existing species.
All organisms currently living on Earth are basically solar powered because, except for a few unique creatures, all life receives energy from the food made by photosynthesis.
Bionote
Redox reactions are common in biological systems. A substance is oxidized when it loses electrons, whereas the substance that gains electrons is reduced. In photosynthesis, water is oxidized and carbon dioxide is reduced.
Photosynthesis
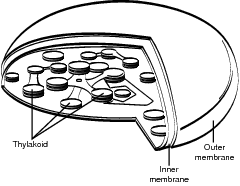
The mechanism for photosynthesis begins with an understanding of the location and structure of the active area in the plant. As previously described, chloroplasts are the organelles where photosynthesis takes place. Within the chloroplasts, the stroma is the location where carbon dioxide is converted into sugar and where the specialized membranes called thylakoids are located. Within the thylakoids, chlorophyll molecules actually trap and process the energy of light with the help of two photosystems described in the following section (“Light Reaction”). Carbon dioxide enters and oxygen exits the plant from adjustable openings on the underside of leaves called stomata. Water is drawn into the plant by its cohesive nature from roots in terrestrial plants and by direct absorption from the environment in aquatic plants. Plants also grow to maximize their exposure to light. Refer to the illustration Chloroplasts.
Photosynthesis is an oxidation-reduction (redox) type of reaction.
Light Reaction
Photosynthesis is divided into two stages. The first stage, called the light reaction, is where the energy of light is captured by the chlorophyll molecules and processed to create high-energy compounds that are used later in the dark reaction (covered in the section that follows). The second stage, known as the Calvin cycle after its discoverer, is also known as the dark reaction, because it uses the energy created in the light reaction to bond carbon chains together to form sugars, other carbohydrates, proteins, lipids, and nucleic acids.
The light reaction occurs in four distinct processes that run continuously if conditions permit:
- Light energy is absorbed by chlorophyll molecules and transferred to make high-energy electrons.
- High-energy electrons enter the electron transport chain where their energy is transferred to electron acceptors.
- Water is oxidized to produce hydrogen ions and the waste gas, oxygen.
- High-energy compounds, ATP and NADH, are formed.
The mechanism for the four processes involves an interaction between structure and function. Within the thylakoid membrane are clusters of pigmented molecules (called photosystems), in addition to chlorophyll, that cooperate to capture and process the energy of light. There are two photosystems that contain 200 to 400 molecules of chlorophyll and other supporting pigments that collectively transfer the light energy to create a high-energy electron(s). Oddly enough, they are called photosystem 1 and photosystem 2, even though photosystem 2 usually begins the reaction.
Bionote
Supporting pigments, such as carotenoids, capture light energy of wavelengths not useful to chlorophyll molecules, as a means of extending the plant's ability to capture more of the energy from the available light. These support pigments usually operate at the lower, less-energetic wavelengths of light, and their colorful presence becomes evident in the fall after the masking chlorophyll molecules are decomposed by the shorter days and cooler weather to expose the orange, yellow, red, and purple pigments that provide the fall colors in deciduous trees (ones that drop their leaves).
When a photon of light hits the photosystem, the pigmented molecules absorb the energy and transfer it to either of two central chlorophyll molecules: P700, which activates photosystem 1; or P680, which activates photosystem 2. P700 and P680 reference two types of chlorophyll molecule. The P stands for “pigment” and the numbers refer to the wavelength of light that activates them.
In the current model, photosystem 2 creates ATP and NADH, both high-energy compounds. Whenever a photon of light is trapped by P680 molecules, they transfer the energy to one of its electrons. This energizes or “excites” the electron to an energy level beyond the attractive force of the nucleus, and it leaves the P680 molecule to immediately be received by an electron-acceptor molecule, which channels its entry into the electron transport chain. The P680 chlorophyll molecule is oxidized and the electron-acceptor molecule is reduced.
The electron transport chain, located in the thylakoid membrane, is a series of molecules that systematically remove energy from the electron as it moves from molecule to molecule. The energy subtracted from the electron is used to move protons (hydrogen ions, H+) into the thylakoid. The extra protons (which were created when water was oxidized) in the thylakoid membrane set up a potential energy gradient, much like water when impounded behind a dam. As the hydrogen ions push and return through channel protein gates in the membrane, these specialized protein gates use that kinetic energy to catalyze a phosphorylation reaction that adds a high-energy phosphate group to an ADP molecule, creating ATP.
Meanwhile, light energy is also absorbed by the P700 chlorophyll molecule in photosystem 1, which also transfers the energy to its electrons, which excites them to enter a different electron transport chain. The oxidized P700 molecule then instantly attracts the loose and energy-rich electrons created in the photosystem 2 electron transport chain to replenish its electron cloud. The excited electron from photosystem 1 combines to reduce NADP+ to the energy-rich compound NADH. Refer to the illustration Photosystem Model.
In summary, within the light reaction, there is a continuous flow of electrons from water to photosystem 2, which creates energy-rich ATP and provides energy-depleted electrons to photosystem 1, which then replaces the excited electrons that enter a different electron transport chain to create NADH. The light reaction harnesses the energy of light and transfers it to the chemical energy of a molecule.
Calvin Cycle or Dark Reaction
The Calvin cycle is called the dark reaction because it does not need light to make biomolecules from the energy created in the light reaction. The Calvin cycle is explained in three steps:
- Formation of PGA, a three-carbon molecule
- Conversion of PGA into PGAL
- Recovery of the starting material and formation of organic compounds
In Step 1, carbon dioxide bonds with the five-carbon RuDP (ribulose diphosphate) molecule to create a temporary six-carbon molecule that immediately splits into two, three-C molecules called PGA.
In Step 2, PGA receives a high-energy phosphate group from ATP (de-energizing ATP to ADP, which can then be reused in the light reaction). Next, NADH adds a proton (hydrogen ion) and releases a phosphate group, thus creating PGAL and the now energy-poor NADP molecule.
In Step 3, most of the newly created PGAL is converted into RuDP, which can then re-enter and restart the Calvin cycle. However, one out of every six PGAL molecules is converted into organic compounds needed elsewhere by the cell.
Statistically, six revolutions of the Calvin cycle are needed along with the addition of six carbon dioxide molecules to create a six-carbon sugar such as glucose. So, technically, the simplistic equation for photosynthesis is correct, however, not explanatory. A better representation is the following:
- Light reaction: Water + ADP + NADP + Phosphate + Light energy → ATP + NADH + Oxygen
- Calvin cycle: ATP + NADPH + RuDP + Carbon dioxide → PGAL + NADP+
Alternative Carbon-Fixing Pathways
It is worth noting that two alternative mechanisms have evolved in response to different environmental conditions for converting carbon dioxide into organic compounds, called carbon fixation: C4 and CAM. The C4 method has unique enzymes that combine inorganic carbon dioxide to create a four-carbon intermediate molecule rather than the three-carbon molecules of the Calvin cycle. The C4 mechanism is an advantage for certain types of plants because on hot, sunny days, C4 plants partially close their stomata to minimize carbon dioxide entry and oxygen release. Normally this greatly reduces the carbon-fixation capabilities; however, C4 plants have an additional enzyme that allows the formation of four-carbon molecules from carbon dioxide even when the carbon dioxide concentrations are low. The four-C molecules then deliver the carbon dioxide to the Calvin cycle, where it is processed. C4 plants, such as crabgrass and corn, only have an advantage during the hot summer months because the energy cost of the C4 mechanism is more expensive than C3 (light reaction).
Bionote
Aerobic cellular respiration is the exact opposite of photosynthesis. The starting products of either reaction are the end product of the other, and vice versa. They are balanced, opposing reactions.
CAM mechanics are designed to conserve water for succulent plants, such as cacti and pineapples, that have adapted to live in hot, dry biomes. Unlike all other plants, CAM plants only open their stomata at night, when they are least likely to lose water by evaporation. Therefore, they take in and fix carbon dioxide at night and create organic compounds that release the carbon dioxide during the day to enter the Calvin cycle. Because they operate at night, their energy-trapping and absorption capacity is limited, so they grow slowly; however, they retain water very well and are therefore highly adapted to these adverse conditions. The sum total of all photosynthetic pathways is the same: the production of glucose (and other organic compounds).
Excerpted from The Complete Idiot's Guide to Biology © 2004 by Glen E. Moulton, Ed.D.. All rights reserved including the right of reproduction in whole or in part in any form. Used by arrangement with Alpha Books, a member of Penguin Group (USA) Inc.
To order this book direct from the publisher, visit the Penguin USA website or call 1-800-253-6476. You can also purchase this book at Amazon.com and Barnes & Noble.
Trending
Here are the facts and trivia that people are buzzing about.





